
生物物理所在III-E型CRISPR-Cas系統研究方面取得進展
軍工資源網 2023年01月30日CRISPR-Cas系統廣泛存在于細菌、古菌和某些細菌的病毒中(bacteriophage),可以特異性識別并降解外源入侵的基因。根據CRISPR-Cas系統干擾機制的不同,這些系統被分為兩類,即Class1和Class2。Class1由多個蛋白構成效應復合物(包括I、III和IV型),Class2由單個蛋白行使功能(包括II、V和VI型)。III型CRISPR-Cas系統進一步可分為III-A和III-B兩個亞型,其特點是具有Cas10蛋白。Cas10通過與Cas5、多個拷貝的Cas11、Cas7以及一條crRNA(CRISPR RNA)組裝成多亞基效應復合物。III-A和III-B型CRISPR-Cas系統的多亞基構成在某種程度上限制了這些系統作為基因編輯和檢測工具的可操作性。
2020年,Makarova等報道了新型的III型CRISPR-Cas系統,即III-E型CRISPR-Cas系統。生物信息學分析顯示該系統的關鍵蛋白(稱為Cas7-11或gRAMP)是一個包含有數個Cas7結構域和一個Cas11結構域的單鏈蛋白。生化實驗進一步表明,Cas7-11能夠加工pre-crRNA使之成為成熟的crRNA,且Cas7-11能夠特異性切割單鏈目標RNA(target RNA,tgRNA)底物。這一生化特點和VI型CRISPR-Cas系統中的Cas13a相似(Class2),但Cas7-11并不顯示出Cas13a所具有的底物旁切活性,且Cas7-11敲低細胞RNA時無明顯的細胞毒性,這使得Cas7-11在被改造成為一款可靠的RNA操控工具方面頗具潛能。Cas7-11基因附近含有多個CRISPR-Cas相關蛋白,包括Csx29(TPR-CHAT)、Csx30、Csx31和RpoE等。尤其是Csx29,可與Cas7-11形成穩定的復合物,并能夠抑制Cas7-11切割tgRNA的活性。Csx29由N端的TPR結構域和C端的CHAT結構域組成。CHAT結構被認為具有可調控的蛋白酶活性且與細胞程序性死亡相關。上述研究揭示了III-E型CRISPR-Cas可能具有全新的免疫應答機制。然而,Cas7-11如何加工pre-crRNA、如何識別并切割tgRNA,Csx29如何調節Cas7-11的活性,Csx29如何被激活等機制尚不清楚。為了回答這些科學問題,中國科學院生物物理研究所對來自Desulfonema ishimotonii菌的DiCas7-11-crRNA復合物開展了結構與功能研究,并解析了一系列該復合物相關的冷凍電鏡結構(圖1)。
該研究解析了DiCas7-11-crRNA未結合底物tgRNA的3.53?的冷凍電鏡結構。結構分析揭示了DiCas7-11-crRNA結構整體呈現“海馬”狀。DiCas7-11由4個Cas7L(Cas7-like)結構域(Cas7L1-4)、1個Cas11L結構域和1個Insertion結構域組成,且一條長38nt的crRNA(15 nt 5' handle + 23 nt spacer)貫穿于整個DiCas7-11蛋白內部。15 nt 5' handle和Cas7Ll、Cas7L2緊密結合,進一步的生化實驗證實Cas7L1保守的殘基His43可切開pre-crRNA的-15和-16堿基之間的磷酸二酯鍵以生成成熟的crRNA。為了剖析DiCas7-11-crRNA如何識別并切割tgRNA,該團隊解析了DiCas7-11-crRNA與tgRNA復合物的3.64?的冷凍電鏡結構。結構分析揭示了crRNA和tgRNA形成堿基互補配對,闡釋了Cas7L2和Cas7L3切割tgRNA的結構細節。刪除Insertion結構域能夠增強DiCas7-11切割tgRNA活性的同時不損害其切割底物的特異性和加工pre-crRNA的能力,這表明可通過刪除Insertion結構域可將DiCas7-11改造成為更小的RNA操控工具。研究進一步解析了DiCas7-11-crRNA和含有與5' handle配對的 3' anti-tag的tgRNA(tgRNA中與crRNA 5' handle對應的部分稱為3' anti-tag)復合物的冷凍電鏡結構(3.46?),以及DiCas7-11-crRNA和含有與5' handle不配對的3' anti-tag的tgRNA復合物的冷凍電鏡結構(3.14?),同時,從結構和生化上揭示了tgRNA的3' anti-tag不參與DiCas7-11-crRNA切割底物活性的調節,這不同于Cas13a的重要性特性。
為了探究DiCsx29調節DiCas7-11切割tgRNA的分子機制,科研人員解析了DiCas7-11-crRNA-DiCsx29復合物3.29?的電鏡結構,發現了DiCas7-11-crRNA結構中兩段觀察不到的區域即FR1(366-397 aa)和FR2(1316-1337 aa)在結合DiCsx29后變得穩定有序。FR1與FR2通過形成一對氫鍵共同阻礙了tgRNA與crRNA在A1至G9的堿基配對,從而調控了DiCas7-11切割tgRNA活性,這一結論隨之得到生化實驗的證實。此外,DiCas7-11-crRNA-Csx29切割tgRNA同樣不依賴5' handle堿基的配對與否。通過DiCas7-11-crRNA-Csx29與III-A型CRISPR-Cas系統以及相關結構的比較,研究推測DiCsx29活性的激活可能依賴tgRNA 3'anti-tag的調控,這一假設得到該研究后續生化實驗的支持。當tgRNA的3' anti-tag與crRNA 5' handle堿基不配對時,DiCsx29蛋白酶活性被激活并切割DiCsx30,被切割后的DiCsx30可能引起宿主的死亡(圖2)。該研究進一步發現,當tgRNA與crRNA 5' handle堿基不配對大于4 nt,且與spacer堿基配對不少于12 nt時,DiCsx29均可以有效切割DiCsx30。由于該蛋白的重要性,近期有多個類似的工作發表,與這些工作相比,研究人員在crRNA的3' 端區域充當“種子區域”(seed region)的作用來調節DiCas7-11活性,以及利用FR1與FR2所形成的“離子鎖”的別構效應來調節DiCas7-11活性等方面提出了較為新穎的觀點(圖2)。
該研究詳細闡述了DiCas7-11加工pre-crRNA和切割tgRNA的機制、DiCsx29對DiCas7-11切割tgRNA活性調控的機制以及DiCsx29蛋白酶活性激活的調控機制。該工作豐富了科學家對CRISPR-Cas系統的認知,有助于將DiCas7-11改造為新型的、無細胞毒性的RNA操控工具。
1月26日,相關研究成果以Cryo-EM structure and protease activity of the type III-E CRISPR-Cas effector為題,發表在Nature Microbiology上。研究工作得到科技部、中科院戰略性先導科技專項的支持。該研究樣品的鑒定、冷凍電鏡樣品的篩查和數據收集工作得到生物物理所蛋白質科學研究平臺和生物成像中心等的幫助。
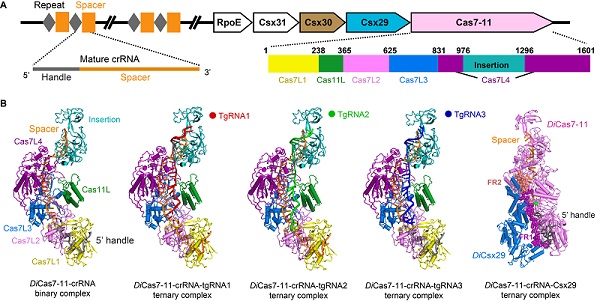
圖1.A、Desulfonema ishimotonii III-E型CRISPR-Cas系統的基因組成形式;B、解析的一系列DiCas7-11-crRNA相關復合物的結構。
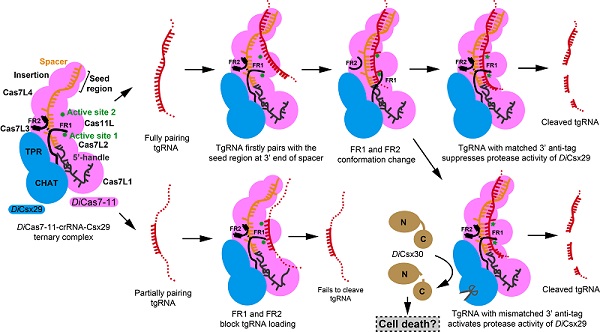
圖2.DiCas7-11-crRNA-Csx29活性的調節